



A tengericsikó meséje II: A gamma és az éleshullám aktivitás keletkezése
Ezzel a kérdéssel fejeztük be: "Hogyan tud ugyanaz a hippokampáis idegsejthálózat több különböző aktivitásmintázatot előállítani?"
Ezt a kérdést vizsgáltuk majd egy évtizedig. Megválaszolásához az in vitro agyszelet technikát használtuk. A hippokampuszból 600 µm (0.6mm) vastag agyszeleteket lehet készítni, melyekben több tízezer sejt őrzi meg kapcsolatrendszerét, és megfelelő körülmények biztosítása esetén működését.
 Az ilyen szeletek lehetővé teszik, hogy mikroszkóp alatt 1-2 vagy 3 elektródával elvezessük egyrészt a hálózat csoportos (populációs) aktivitását mutató EEG jeleket, másrészt egyedi idegsejtek aktivitását. Hájos Norbert kollegám kidolgozott egy olyan szeletfenntartó kamrát, amelyben a kis hippokampusz darabok, a fenntartó oldathoz adagolt kéreg alatti átvivő anyag analóg (charbachol, mely az acetil-kolin hatását utánozza) jelenlétében gamma aktivitást, annak hiányában SWR-ket állítottak elő. Megvolt tehát egy modell rendszerünk arra, hogy azt vizsgáljuk, hogyan áll elő a hippokampusz serkentő és gátló sejtjeinek kölcsönhatásából a két aktivitásmintázat.
Az ilyen szeletek lehetővé teszik, hogy mikroszkóp alatt 1-2 vagy 3 elektródával elvezessük egyrészt a hálózat csoportos (populációs) aktivitását mutató EEG jeleket, másrészt egyedi idegsejtek aktivitását. Hájos Norbert kollegám kidolgozott egy olyan szeletfenntartó kamrát, amelyben a kis hippokampusz darabok, a fenntartó oldathoz adagolt kéreg alatti átvivő anyag analóg (charbachol, mely az acetil-kolin hatását utánozza) jelenlétében gamma aktivitást, annak hiányában SWR-ket állítottak elő. Megvolt tehát egy modell rendszerünk arra, hogy azt vizsgáljuk, hogyan áll elő a hippokampusz serkentő és gátló sejtjeinek kölcsönhatásából a két aktivitásmintázat.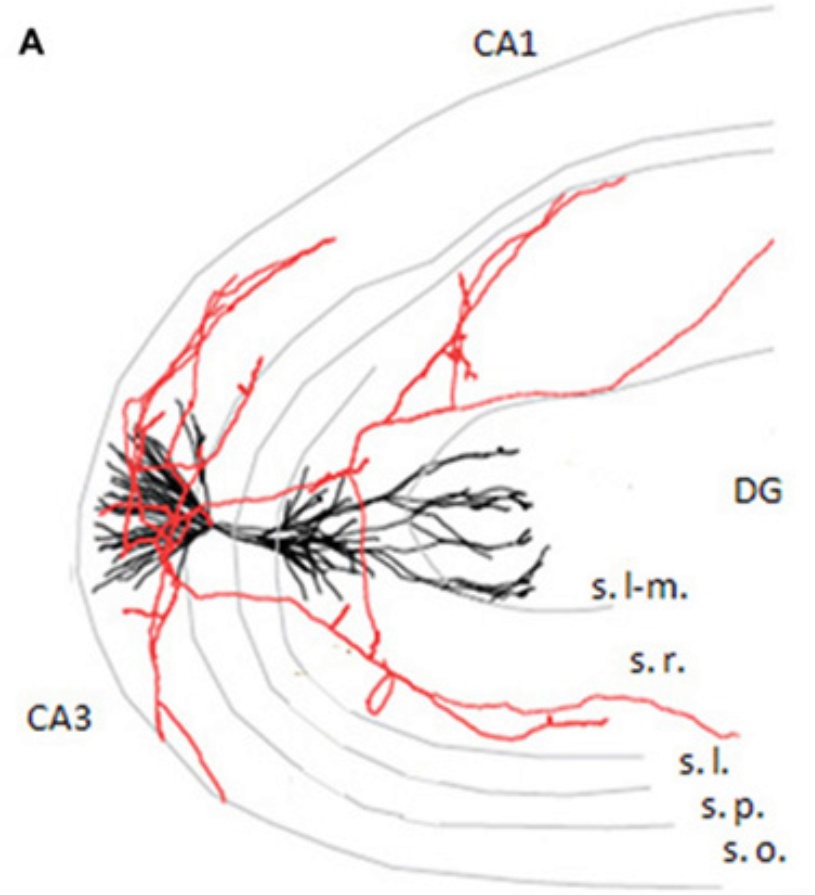 Hogy a részleteket bemutathassuk, el kell merülnünk egy kicsit a CA3 terület anatómiájában, ugyanis itt dolgoztunk, mert élő állatokból származó EEG kísérletek alapján, ez a terület a kiindulópontja mind a gamma, mind az éleshullám aktivitásnak. Mint írtuk, ennek a területnek a jellemzője az, hogy két tetszőleges serkentő sejtje viszonylag nagy valószínúséggel van összekötve, azaz a CA3 hálózat erősen visszacsatolt, rekurrens. Míg ez a valószínúség a hippokampusz CA1 rétegében 0.1%, azaz 1:1000, addig ez a CA3-ban 2-3%, azaz 1:30-1:50. Ezenkívül hasonlóan erős kapcsolat található a serkentő sejtekről a gátlósejtek felé. Ennél még erősebb a gátlósejtekről a serkentő sejtekre érkező gátlás. Ennak valószínűsége 100%, azaz egy gátlósejt a környezetében található összes serkentő sejtet beidegzi. Ráadásul korábbi munkáink alapján többszörös kapcsolattal. Igaz ez a sejttesten végződő kosársejtekre és a dendriteken végződő sejtekre is. A CA3 tehát tartalmaz egy erősen visszacsatolt serkentő hálózatot, melyet szerencsére egy igen hatékonyan csatolt gátlórendszer tart ellenőrzése alatt.
Hogy a részleteket bemutathassuk, el kell merülnünk egy kicsit a CA3 terület anatómiájában, ugyanis itt dolgoztunk, mert élő állatokból származó EEG kísérletek alapján, ez a terület a kiindulópontja mind a gamma, mind az éleshullám aktivitásnak. Mint írtuk, ennek a területnek a jellemzője az, hogy két tetszőleges serkentő sejtje viszonylag nagy valószínúséggel van összekötve, azaz a CA3 hálózat erősen visszacsatolt, rekurrens. Míg ez a valószínúség a hippokampusz CA1 rétegében 0.1%, azaz 1:1000, addig ez a CA3-ban 2-3%, azaz 1:30-1:50. Ezenkívül hasonlóan erős kapcsolat található a serkentő sejtekről a gátlósejtek felé. Ennél még erősebb a gátlósejtekről a serkentő sejtekre érkező gátlás. Ennak valószínűsége 100%, azaz egy gátlósejt a környezetében található összes serkentő sejtet beidegzi. Ráadásul korábbi munkáink alapján többszörös kapcsolattal. Igaz ez a sejttesten végződő kosársejtekre és a dendriteken végződő sejtekre is. A CA3 tehát tartalmaz egy erősen visszacsatolt serkentő hálózatot, melyet szerencsére egy igen hatékonyan csatolt gátlórendszer tart ellenőrzése alatt.
 |
 |
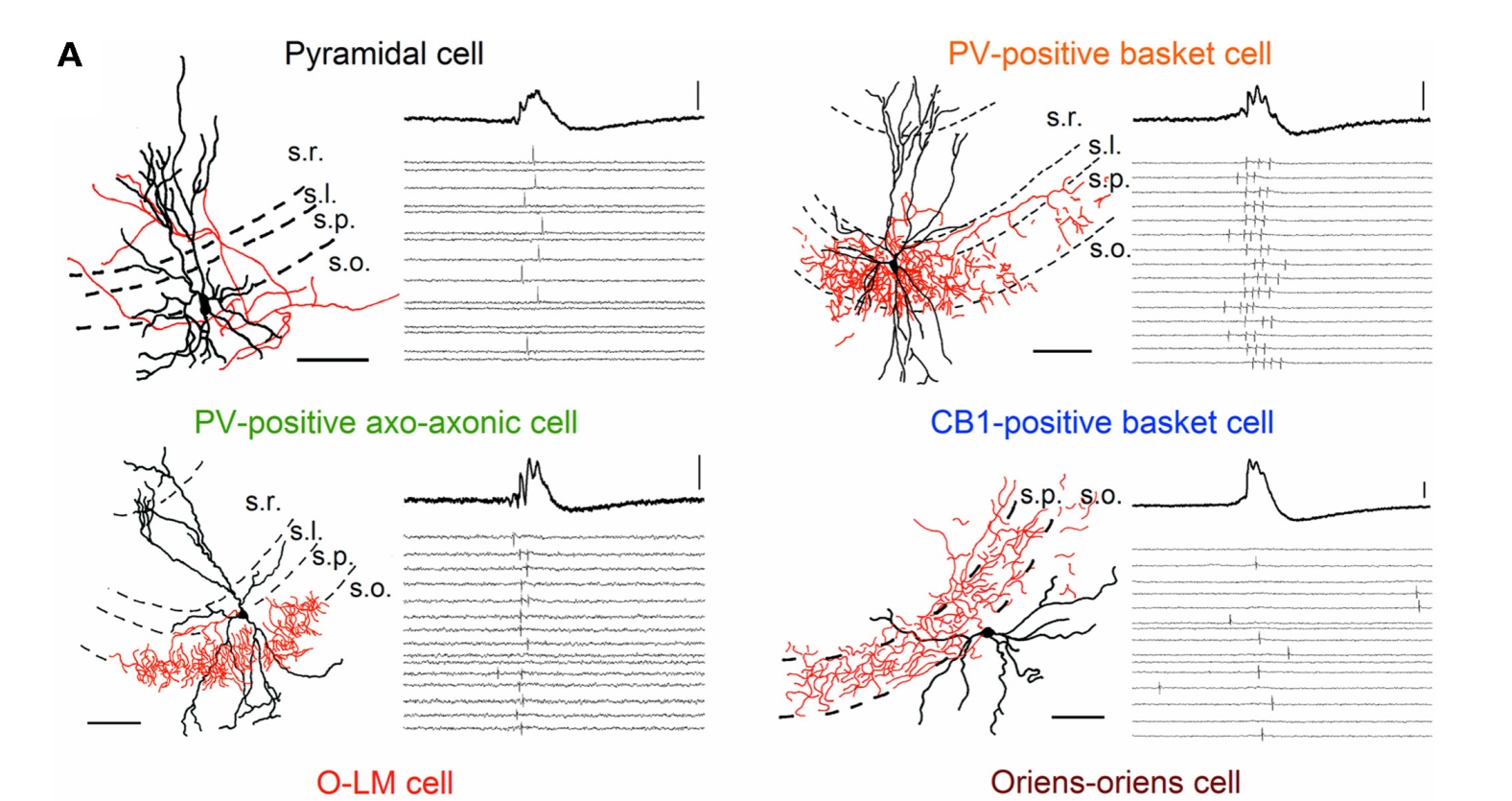
Hogyan is álltunk neki? Először is megnéztük azt, hogy a CA3 serkentő piramissejtjei és számos eltérő gátlósejt csoportja a háttér (gamma és SWR) aktivitás mely fázisaiban tüzel. Ehhez sok száz szelet sok száz anatómiailag azonosított sejtjéből kellett jeleket elvezetni, és összehasonlítani a hálózat EEG aktivitásával, mind gamma ,mind SWR aktivitás alatt. Ebből megtudtuk azt, hogy az eltérő sejtek időben hogyan aktiválódnak, ki-ki előtt és után tüzel, azaz a serkentősejtek milyen gátlósejteket serkentenek és milyen gátlósejtek mikor fejtik ki gátlásukat, mely a serkentősejtek jelintegrációját befolyásolja.
 Kicsit olyan mint a szociológus munkája, aki mondjuk a metró bejáratánál ül és különböző napszakokban megkérdezi az arrajárókat, hogy mi a munkájuk. Kora hajnalban pékeket és buszvezetőket fog fogni, akik munkájukba igyekeznek. Reggel van a legnagyobb forgalom, ilyenkor járnak a nappali műszakosok munkába. Délelőtt nyugi van, de a délutáni csúcsba a hazatérők mellé keverednek az esti műszakosok, portások, biztonsági őrök, takarítók. És így tovább. Ebből aztán ha még nem értené, összerakhatná, hogy a munkát a pékek és buszvezetők indítják és az éjjeli őrök meg takarítók zárják, a többiek középen szorgoskodnak.
Kicsit olyan mint a szociológus munkája, aki mondjuk a metró bejáratánál ül és különböző napszakokban megkérdezi az arrajárókat, hogy mi a munkájuk. Kora hajnalban pékeket és buszvezetőket fog fogni, akik munkájukba igyekeznek. Reggel van a legnagyobb forgalom, ilyenkor járnak a nappali műszakosok munkába. Délelőtt nyugi van, de a délutáni csúcsba a hazatérők mellé keverednek az esti műszakosok, portások, biztonsági őrök, takarítók. És így tovább. Ebből aztán ha még nem értené, összerakhatná, hogy a munkát a pékek és buszvezetők indítják és az éjjeli őrök meg takarítók zárják, a többiek középen szorgoskodnak.
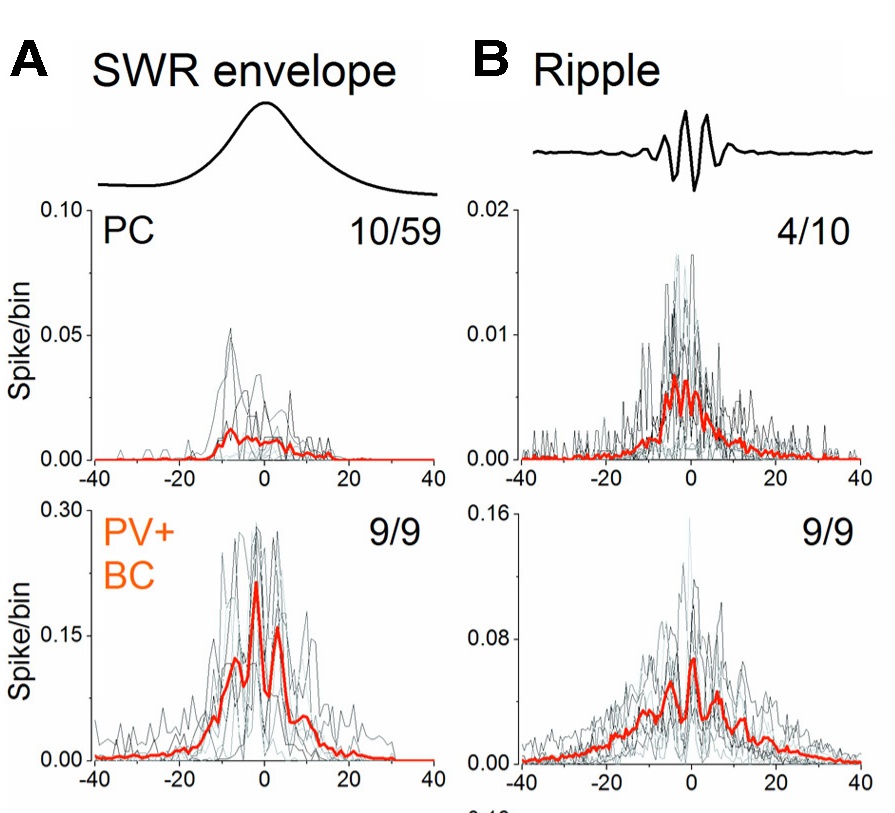
Mit tudtunk meg mindebből? A gamma és az SWR alatt más a sejtek kölcsönhatása, máshogy működnek ugyanazok a sejttípusok, pedig összeköttetés-hálózatuk szerkezete nem változik meg .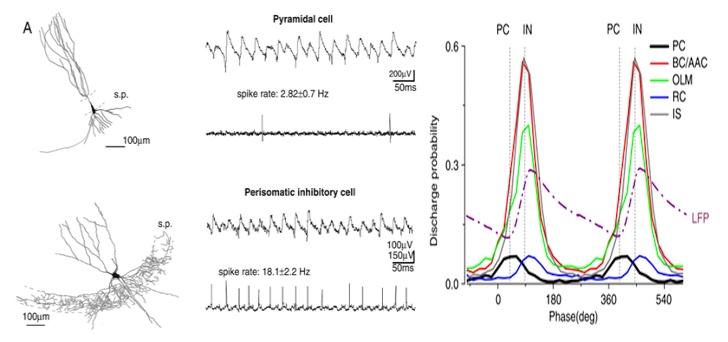
A gamma aktivitás ciklusa azzal kezdődik, hogy a serkentősejtek elkezdenek tüzelni. Amikor aktivitásuk elér egy bizonyos szintet, a parvalbumin (PV) tartalmú kosársejtek, melyek a sejttesten fejtik ki gátlásukat és ezért a leghatékonyabb gátlást -a tüzelés megállítását- okozzák, elkezdenek viszonylag egyszerre kisülni és megállítják a serkentősejtek tüzelését. 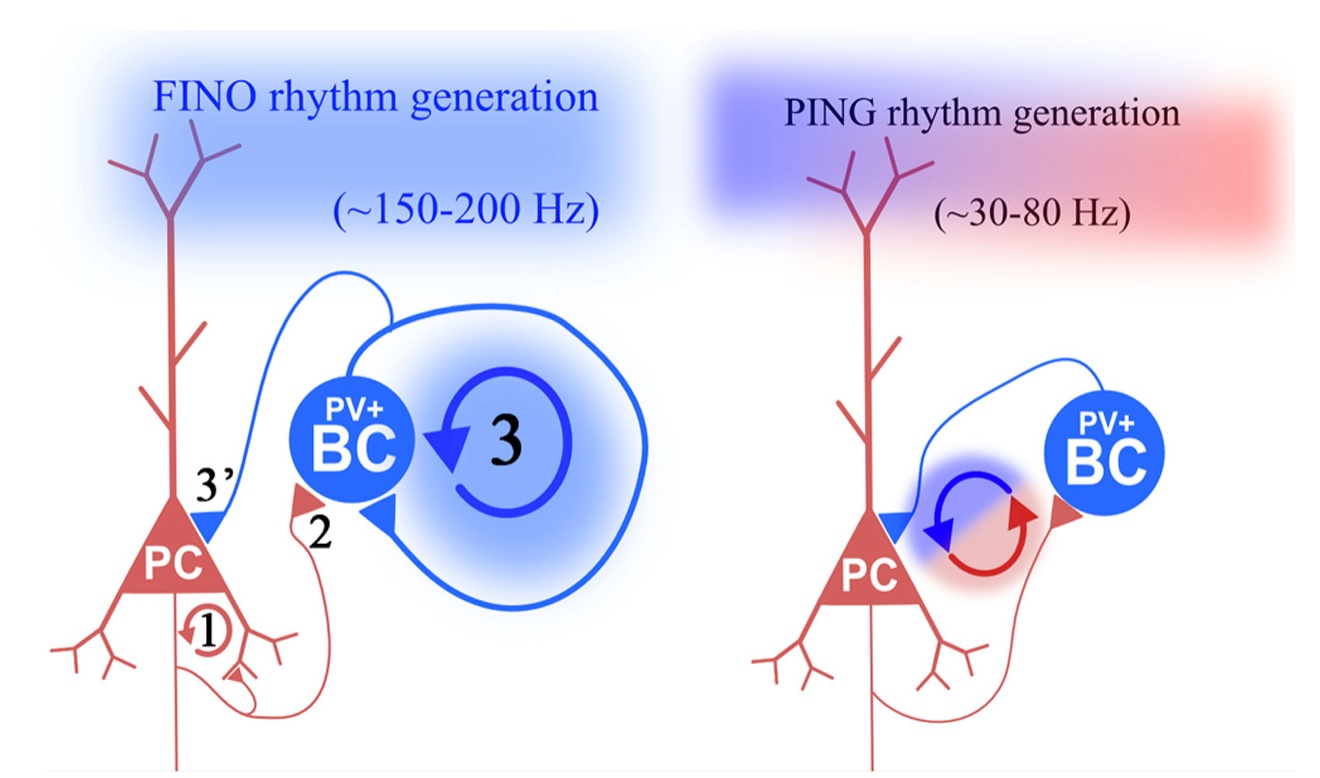 Amikor aztán elmúlik a gátlás hatása (4-8 msec) a serkentősejtek elkezdenek újra tüzelni. Ez a hintázás másodpercenként 40-szer megimétlődik (25 msec-enként) és az összehangolt gátló áramok adják az EEG-ben megfigyelhető gamma hullámokat (melyek 40Hz el érkeznek).
Amikor aztán elmúlik a gátlás hatása (4-8 msec) a serkentősejtek elkezdenek újra tüzelni. Ez a hintázás másodpercenként 40-szer megimétlődik (25 msec-enként) és az összehangolt gátló áramok adják az EEG-ben megfigyelhető gamma hullámokat (melyek 40Hz el érkeznek).
SWR-k alatt egészen más történik (hogy miért arról hamarosan). Ilyenkor az agyszeletek átlagban jelentősen csendesebbek, mint gamma aktivitás alatt. Néha 1-2 másodpercig néhány pirami sejt kivételével alig szólal meg valaki. Aztán egyszercsak, körülbelül 1/10-ed másodperc alatt, a serkentősejtek aktivitása elkezd meredeken nőni, egyenlőre még nem egymáshoz rendezve. Ez eredményezi az éles hullám lassú burkoló görbéjét. Amikor aztán a serkentő hálózatban az aktivitás elér egy küszöböt (mint a gammánál), a PV tartalmú kosársejtek elkezdenek kisülni, de ilyenkor a gyorsan felépülő erős serkentés miatt egymáshoz hangoltan, nagy frekvenciával ismételten sülnek ki. Ez az összehangolt gátlás a serkentősejteket is összehangolt kisülésre készteti. A két sejttípus kisülése és a gyors gátló áramok adják az éleshullám csúcsán a 160Hz-es fodor (ripple) oszcillációt. 4-8 összehangolt kisülés után aztán a serkentősejtek kifáradnak, az SWR elhal, és hosszú ideig csend honol a szeletben, mindaddig míg a kipihent serkentősejtek egy újabb serkentő sejt kisülés lavinát, újabb SWR-t indítanak el.
Szerző: Gulyás Attila