



Different mechanisms in the storage of spatial memory
An extraordinary report on an extraordinary observation! How do the place cells of the hippocampus work? The Nusser group's discovery was published in the journal eLife and the first author Máté Sümegi reports on it.
- A photograph in which my hands are blurred as I flail about. I am about to fall on my stomach. As a memory, it is amusing, but not very instructive. However, if it had been taken with an action camera, unlike a video, we would now be able to see what happened in the seconds before and after the shutter button was pressed. It is a useful technology if we want to capture both the cause and the effect.
The previous example highlights one of the weaknesses of the classic Hebbian learning model. According to this model, learning only occurs through the millisecond timing of nerve cell activity and repeated repetition. If this is true, then how can one-time experiences such as seeing a bear and being slapped be learned?
This highlights the significance of a study published in 2017 by Jeff Magee's laboratory (Jenilia Farm)1, which describes for the first time a synaptic learning mode corresponding to the behavioral time scale of living organisms (behavior time scale synaptic plasticity, BTSP) in the case of hippocampal place memory cells that record spatial position. Previously, another study2 from the same research institute, Albert Lee's lab, presented the role of synaptic learning without BTSP (non-BTSP) in the formation of place memory.
Both observations are fundamental and indisputable. However, due to the technological level of the time, they are based on only a few observed cases. We investigated the contradiction between them using modern technology capable of recording the activity of thousands of hippocampal cells over several consecutive days. Using genetic tools and advanced microscopy, we visualized the intracellular calcium ion level fluctuations accompanying nerve cell activity while mice performed orientation tasks in a virtual reality system. Using computational tools, we were able to record and classify thousands of events in which a hippocampal pyramidal cell suddenly became a place memory cell or added a new location to its list of familiar places, as indicated by increased activity.
What do the results show? How are cell-level place memories ultimately created? The answer is unique: both!
After 163 navigation experiments with 45 animals and the observation and rigorous classification of 22,325 place memory cells, we observed 311 BTSP-based and 58 non-BTSP-based learning events while the mice walked through a familiar corridor.
During these experiments, we also observed a rare but very striking phenomenon. Sometimes, from one circuit to another, an animal's place-specific neural activity pattern (i.e., its spatial representation) in the hippocampus changed dramatically without the animal getting lost. Such neuronal remapping was characterized by a higher than usual rate of place memory formation; in just a few such events, we recorded 40 BTSP and 22 non-BTSP events.
In another experiment, we opened a new section of the corridor to the animals. In this case, the ratio of BTSP to non-BTSP events changed. The 43/26 ratio indicates a relative increase in non-BTSP events due to the novelty effect.
A characteristic feature of place memory formed with BTSP is that the size of the covered area depends on the speed of travel at the time of recording. Similarly, an action camera records a longer distance during the exposure period when we move faster. This feature was not characteristic of non-BTSP-based memory traces, which retrospectively confirms that our selection method did indeed distinguish between two different modes of memory recording.
The formation of BTSP-based spatial memory in the cell plasma always begins with a large and spectacular peak in calcium ion concentration. We have already seen that it is possible to form spatial memory at the cellular level (non-BTSP) even without such large peaks. We were curious to know whether such high calcium activities are sufficient to form spatial memory. We were able to present 59 examples in which a pyramidal cell recorded a location in its memory repertoire using a BTSP mechanism that began with a large calcium spike. Meanwhile, calcium spikes of at least the same magnitude or even greater did not result in the encoding of new place memories in the same cell. In other words, taking technical limitations into account, we can say that high calcium ion concentration spikes alone are not sufficient for BTSP-based place memory encoding.
In summary, both BTSP and non-BTSP mechanisms play a role in the storage of place memory in hippocampal pyramidal cells. Their contribution ratio depends on the context and on the remapping events observed in an unchanged environment. The high peak concentration of calcium ions in the cell body accompanying the BTSP mechanism is not only unnecessary but also insufficient for the formation of spatial memory.
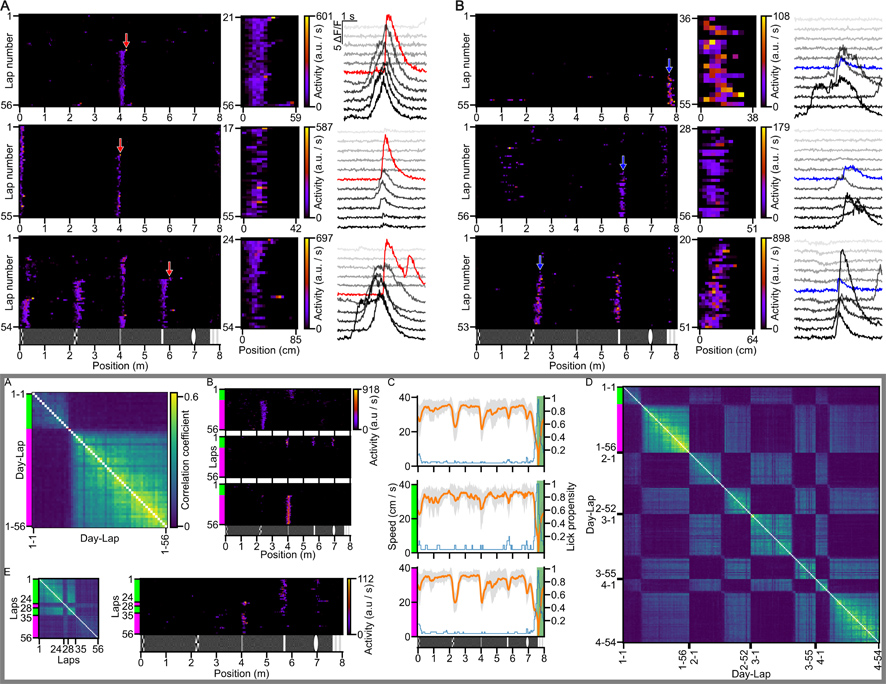
A,B: Examples of BTSP (A) and non-BTSP (B) based place representations.
Boxed section A-E: Examples of neural remapping.
References:
1. Bittner et al., Science, 2017
2. Lee et al., Science, 2012