



What happens in the brain region called the hippocampus while we learn and adapt?
The study, published in Science Advances and resulting from collaboration between Makara and the Ujfalussy group, revealed how the activity patterns of nerve cells in the hippocampus, the brain's main learning and memory center, change during spatial learning and adaptation to sudden changes in the environment. The results show that the hippocampus plays an important role not only in learning but also in brain processes responsible for cognitive flexibility. Further investigation of these processes may lead to a better understanding of the mechanisms underlying cognitive decline observed in many psychiatric disorders and in aging.
One of the most important abilities of our brain, essential for the survival of the species, is to facilitate adaptation to a changing environment. This is based on learning through experience. At the same time, flexible thinking, also known as cognitive flexibility, is necessary to enable rapid adaptation to unexpected environmental changes. The underperformance of the mechanisms that lead to flexible thinking leads to rigid, narrow thinking, or cognitive rigidity. Such patients are able to recognize and learn rules, but when facts and rules change, they cling to previously proven solutions.
Rita Nyilas, the first author of the article, had this to say about why we need to learn more about these processes: "It is not fully understood how the spatial and contextual information necessary for adaptation is incorporated into the brain's cognitive map during experience-based learning. We also know little about the changes that occur during a rule or context change in the hippocampus, the area of the brain that plays a central role in memory processes and navigation.
Our goal was to create an experimental paradigm in which we could simultaneously examine the activity of excitatory cells located in the output region of the hippocampus in mice during context-dependent learning and spatial orientation."
The experiments used transgenic mice that expressed an activity-dependent fluorescent protein (GCaMP) in the target cells to be studied, allowing long-term monitoring of cell activity using two-photon microscopy. The mice had to learn which colors and patterns in their environment corresponded to rewards by running through virtual corridors projected onto them. Two different types of corridors with different walls were used in the experiment and only in one of them did the mouse receive a reward drop if it searched in the right place.
There were several methodological requirements for conducting the experiments. One was the development of details that would enable stable, high-quality imaging over a long period of time (up to several weeks or months), and the other was the optimization of the task performed in the virtual environment. The length of the projected corridor had to be adjusted and the number of experiments recorded per day had to be determined in order to ensure that the animals were motivated and performed the task enough times for statistical analysis of the results, and develop analytical methods. The carefully designed experiments led to several surprising results.
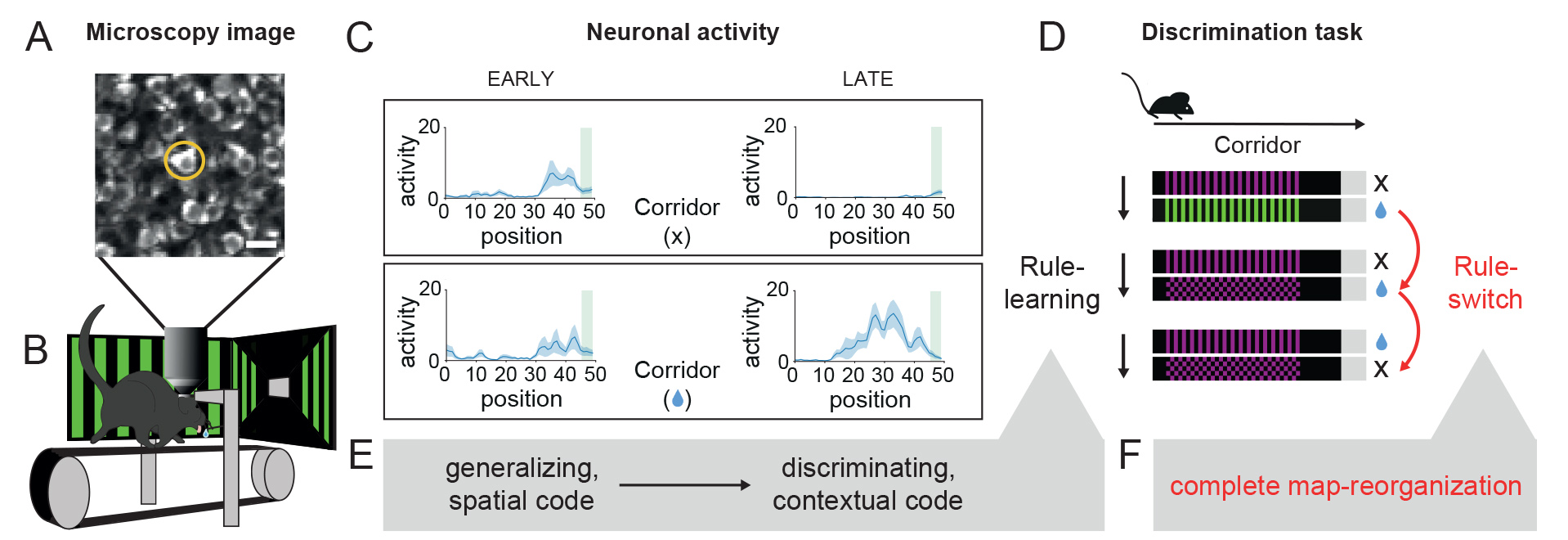
We used two-photon microscopy (A) in mice navigating in virtual corridors (B) to record neuronal activity from the hippocampus (C). Our results show that during learning (E) to discriminate (Rule-learning) between two visually distinct corridors (D), neurons initially generalize between the two environments coding only spatial position (C, EARLY), but then develop a discriminating contextual code with experience (C, LATE). Uncued change of task conditions (Rule-switch) leads to global reorganization of the cognitive map (F).
Rita Nyilas: "When learning the discrimination task we used, spatial coding of corridors first develops in the hippocampus. However, parallel to learning reflected in behavior, the more general spatial representation was gradually supplemented by a corridor-specific, discriminative pattern of neural activity. Surprisingly, cells showing selective activity also appeared in the representation of the non-rewarded corridor, meaning that an abstract, context-dependent code developed in the hippocampus.
When the color and pattern of one of the corridors were unexpectedly changed, i.e., when the context of the already learned task was suddenly altered, the neural code was rearranged very quickly in line with the rapid decline in performance of the mice, but not only in the hippocampal representation of the new corridor, but also in that of the unchanged corridor. This suggests a hierarchical neural mapping of the task, characterized by the rule at the top level, followed by the corridor and then the position. When the rule changes, the other two codes, which are present together in the activity of the cells, also change.
In our third type of experiment, we did not change the type of corridor pair used for the already learned task, but unexpectedly and without warning moved the reward from one corridor to the other. The mice were slower to notice this change in context, so the change in representation also took longer to occur. In this case, too, the activity pattern of the hippocampal neurons changed in line with the behavioral change.
The correlation between behavior and hippocampal neural activity was well demonstrated by the fact that after the reward was moved, when the mice tried in vain to find water in the corridor that had been rewarded before the change, the activity pattern of the nerve cells was more similar to the representation before the change than during the correct searches with the changed strategy."
The results therefore suggest that cognitive rigidity and the inability to adapt to new circumstances are caused by well-defined neural processes. This raises the possibility that the neural processes underlying cognitive decline could also be investigated in the future.