




Különböző mechanizmusok a helymemória tárolásában
Nem mindennapi beszámoló egy nem mindennapi megfigyelésről! Milyen mechanizmus szerint működnek a hippokampusz a helymemóriasejtjei? A Nusser csoport felfedezése az eLife folyóiratban jelent meg és Sümegi Máté első szerző számol be róla.
- Egy fénykép, amin elmosódva kalimpálnak a kezeim. Éppen hasra esem. Mint emlék, mulatságos, de nem túl tanulságos. Bezzeg, ha akciókamerával készült volna, egy videóval ellentétben, most láthatnánk, mi történt az expozíciós gomb lenyomását megelőző és az azt követő másodpercekben. Hasznos technológia, ha meg akarjuk örökíteni a kiváltó okot és az okozatot is.
Az előbbi példa rámutat a klasszikus Hebb-féle tanulási modell egyik gyenge pontjára. E modell szerint a tanulás csak az idegsejtek aktivitásának milliszekundumos időzítése mellett, sokszoros ismétlődés hatására alakul ki. Ha ez igaz, akkor hogyan tanulhatók meg olyan egyszeri tapasztalatok, mint a medve megpillantása és a pofon közötti asszociáció?!
Ez adja a jelentőségét annak a 2017-ben Jeff Magee laborjából (Jenilia Farm) megjelent tanulmánynak1, ami először ír le egy, az élőlények viselkedési időskálájának megfelelő szinaptikus tanulási módot (behavior time scale synaptic plasticity BTSP), a térbeli pozíciót rögzítő hippokampális helymemória-sejtek esetében. Korábban, ugyanabból a kutatóintézetből, Albert Lee laborjából kijött egy másik tanulmány is2, ami a helymemória kialakulásában a BTSP nélküli, szinaptikus tanulás (nem BTSP) szerepét mutatta be.
Mindkét megfigyelés alapvető jelentőségű és megkérdőjelezhetetlen. A kor technológiai szintjéből adódóan azonban csak néhány megfigyelt esetre alapoz. A köztük feszülő ellentmondásnak jártunk utána egy modern, egyszerre ezres nagyságrendű hippokampális sejt aktivitását akár több egymás utáni napon át is rögzíteni képes technológiával. Genetikai eszközökkel és fejlett mikroszkópiával az idegsejtek aktivitását kísérő sejten belüli kálciumionszint-ingadozást tettük láthatóvá, miközben az egerek virtuális valóság rendszerben tájékozódási feladatot végeztek. Komputációs eszközökkel lehetőségünk volt több ezer olyan eseményt rögzíteni és osztályozni, amikor egy hippokampális piramissejt egyik pillanatról a másikra helymemória-sejtté válik, vagy egy új helyszínt vesz fel az általa emelkedett aktivitással jelzett ismerős helyek közé.
Mit mutatnak az eredmények, melyik módon születnek végül a sejtszintű helyemlékek? A válasz különleges: Mind a kettő!
45 állat 163 navigációs kísérlete és 22325 helymemóriasejt megfigyelése és szigorú osztályozása után 311 BTSP és 58 nem BTSP alapú tanulási eseményt figyeltünk meg, miközben az egerek egy megszokott folyosót jártak be.
Ezen kísérletek alatt megfigyeltünk egy ritka, de igen látványos jelenséget is. Néha egyik körről a másikra nagymértékben megváltozott egy állat helyspecifikus idegsejt aktivitásmintázata (azaz térreprezentációja) a hippokampuszban anélkül, hogy eltévedt volna. Az ilyen neuronális újratérképezéseket a szokásosnál is magasabb helymemória-születési arány jellemezte; mindössze néhány ilyen esemény során 40 BTSP és 22 nem BTSP eseményt rögzítettünk.
Egy másik kísérletben egy új folyosórészletet nyitottunk meg az állatok előtt. Ilyenkor a BTSP és nem BTSP események aránya megváltozott. A 43/26-os arány a nem BTSP viszonylagos megszaporodását jelzi az újdonság hatására.
A BTSP-vel formálódó helymemóriára jellemző, hogy a lefedett hely nagysága függ a rögzítéskori haladási sebességtől. Hasonlóan ahhoz, ahogy az akciókamera is hosszabb utat vesz fel az expozíció körüli időszakban, ha gyorsabban haladunk. Ez a tulajdonság nem volt jellemző a nem BTSP alapú memórianyomokra, ami utólag hitelesíti, hogy válogatási módszerünkkel tényleg két különböző memóriarögzítési módot különítettünk el.
A BTSP alapú helymemória kialakulása a sejtplazmában mindig egy nagy és látványos kálciumion-koncentráció csúccsal kezdődik. Azt már láttuk, hogy ilyen nagy csúcsok nélkül is lehetséges sejtszintű helymemória kialakítása (nem BTSP). Kíváncsiak voltunk, hogy az ilyen nagy kálcium-aktivitások elegendőek-e ahhoz, hogy helymemóriát alakítsanak ki. 59 olyan példát sikerült bemutatnunk, amikor egy piramissejt nagy kálcium csúccsal kezdődő BTSP mechanizmussal felvett egy helyet az emlék repertoárjába. Mindeközben legalább ekkora vagy még ennél is nagyobb kálcium csúcsok nem eredményezték új helymemória rögzülését ugyanabban a sejtben. Vagyis a technikai limitációk figyelembevétele mellett elmondhatjuk, hogy a nagy kálciumion koncentráció csúcsok önmagukban nem elegendőek a BTSP alapú helymemória rögzítéséhez.
Összefoglalva, a hippokampális piramissejtek helymemória-tárolásában a BTSP és a nem BTSP mechanizmus is szerepet játszik. Ezek hozzájárulási aránya a kontextustól, illetve a változatlan környezetben megfigyelt újratérképezési eseményektől is függ. A helymemória kialakításához nemcsak nem szükséges, de nem is elégséges a BTSP mechanizmust kísérő magas sejttesti kálciumion koncentráció csúcs.
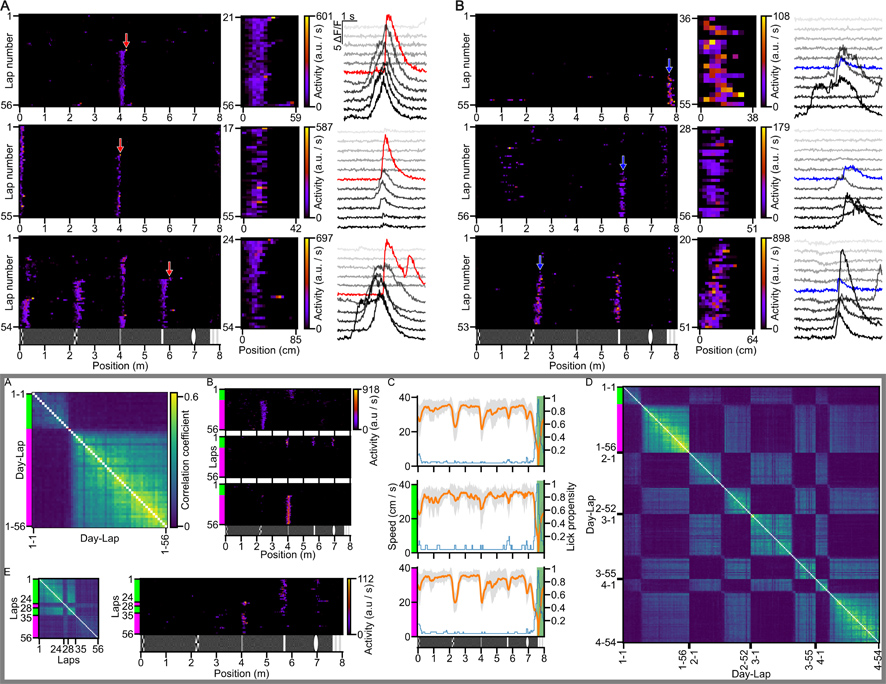
A,B: Példák BTSP (A) és nem BTSP (B) alapú helyreprezentációk születésére.
Keretes rész A-E: Példák a neuronális újratérképezésre.
Hivatkozások:
1. Bittner et al., Science, 2017
2. Lee et al., Science, 2012